Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-07
-
Xenobiotic metabolism primarily occurs in liver which contai
2019-09-19
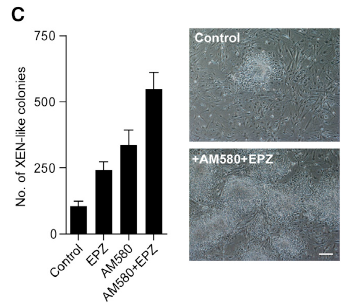
Xenobiotic metabolism primarily occurs in liver, which contains many enzymes to catalyze the transformation of xenobiotic. Usually, cytochrome P-450s (CYP450s) act as the first response for biotransformation of xenobiotic in organisms. CYP450s generally consist of various subfamily enzymes which can
-
br Materials and methods br Results
2019-09-18
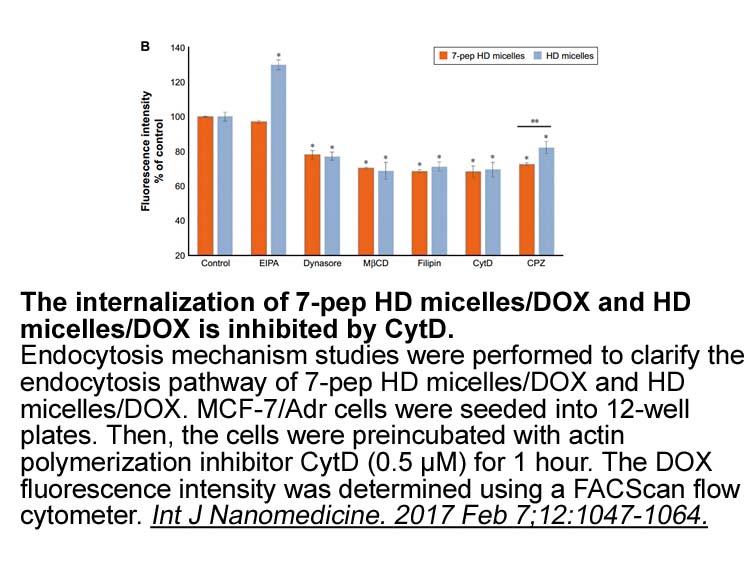
Materials and methods Results Discussion Consistent with previous reports (Barr et al., 2010, Vuong et al., 2010), the current study demonstrates that rats treated with amphetamine exhibit heightened anxiety states during withdrawal. Furthermore, icv. infusion of a CRF2 receptor antagonist
-
Recently several Pt IV complexes
2019-09-18
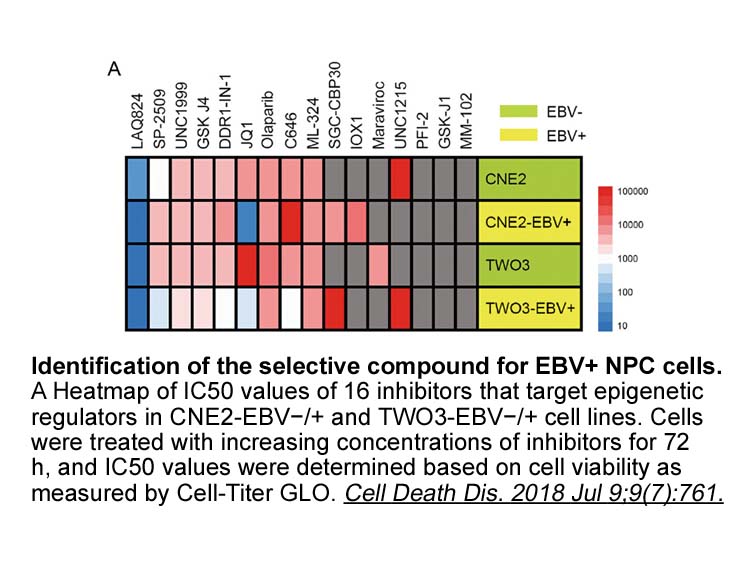
Recently, several Pt(IV) complexes as prodrugs have been designed and studied to enhance anti-tumor efficacy of traditional Pt(II) drugs, improving their cellular uptake and blood-circulation time [26], [27], [28], [29]. Compared with their Pt(II) counterparts, six-coordinated Pt(IV) complexes with
-
Table presents results for estimation of Eq
2019-09-17
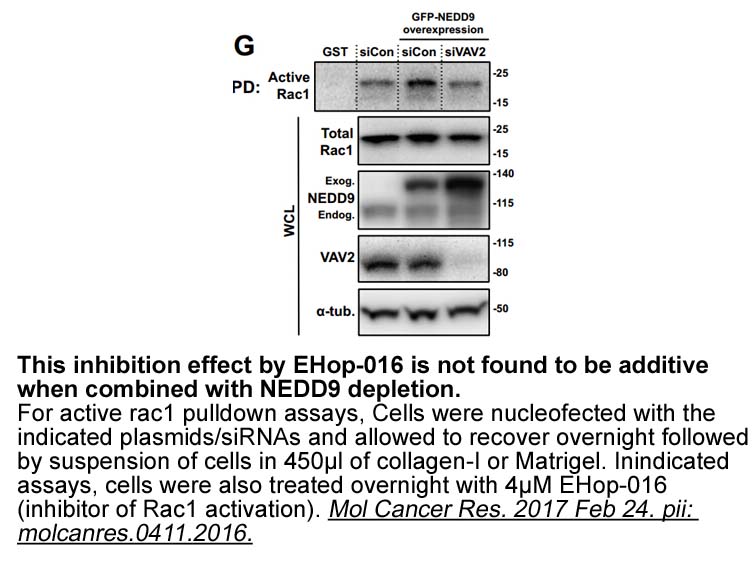
Table 4 presents results for estimation of Eq. (1) using ACTR and SUWE to compare the incidence of attempting the exam between programs. We find alumni are more likely to attempt the exam if they our website do not hold another professional certification, have worked for an accounting firm, have a h
-
5402 Lavallee et al reported that PREG could
2019-09-17
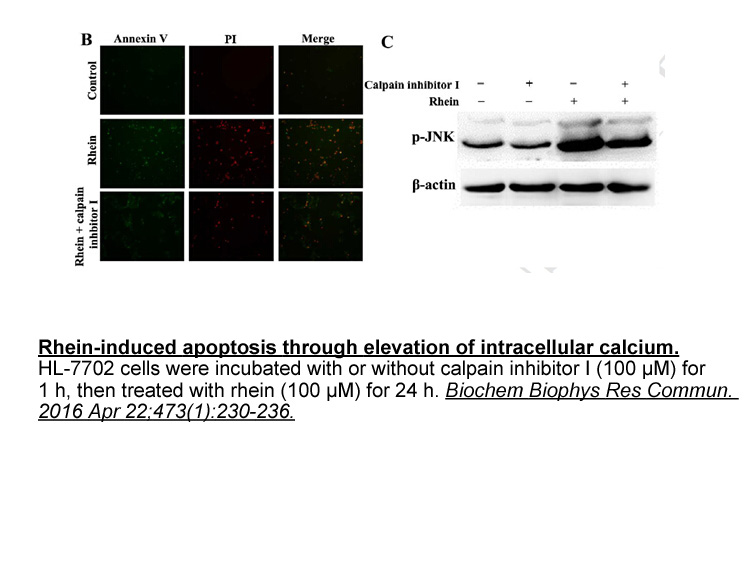
Lavallee et al. [30] reported that PREG could be esterified by lecithin:cholesterol acyltransferase (LCAT). LCAT uses cholesterol and phosphatidyl choline present in the newly formed high-density lipoprotein (HDL) as substrates and convert them into cholesteryl esters and lysophosphatidyl choline. T
-
E ubiquitin ligases are key players in the
2019-09-17
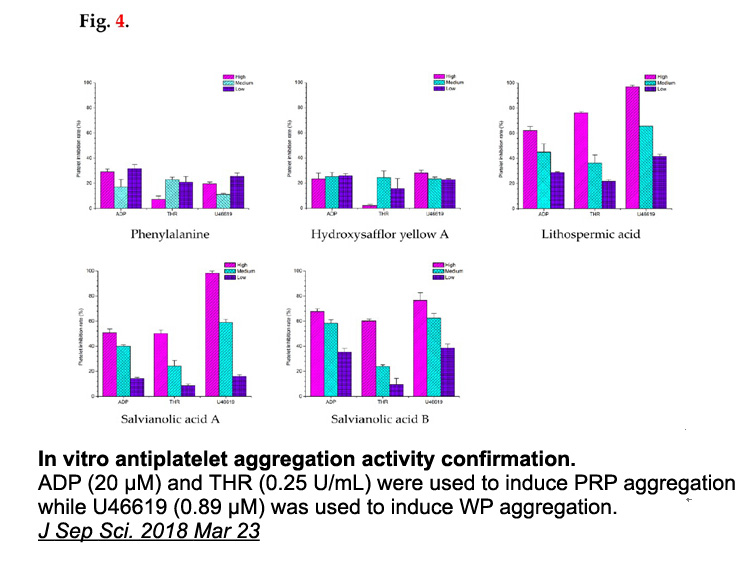
E3 ubiquitin ligases are key players in the ubiquitin-proteasome pathway because they catalyse ubiquitination of substrate proteins.39, 40, 41 As important regulators of cellular ubiquitination, E3 ligases are emerging as attractive drug targets, particularly in cancer.42, 43, 44 However, E3 ligases
-
br Patients and Methods Patients who had been
2019-09-17
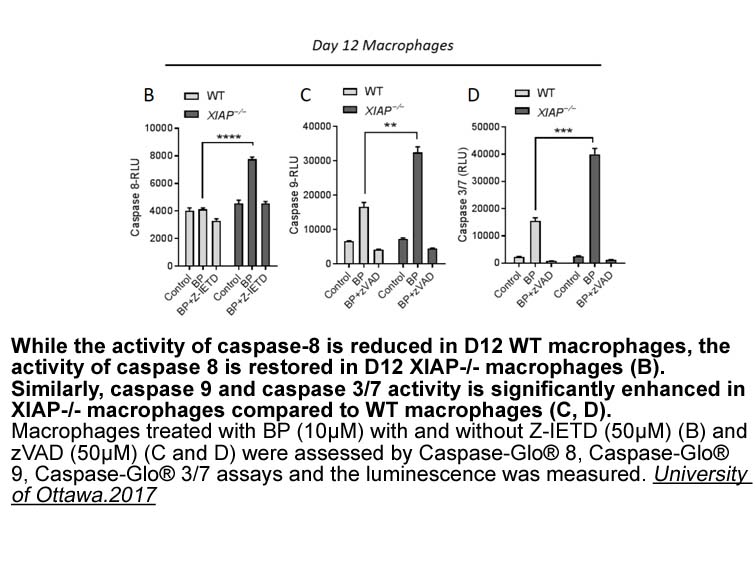
Patients and Methods Patients who had been initiated on a DPP-4 inhibitor (see in the online version at doi:10.1016/j.clinthera.2018.06.002) between April 2012 and June 2017, when the common threshold for dose adjustment for all non-linagliptin DPP-4 inhibitors was a creatinine clearance (CrCl)
-
br DGKs bind and regulate
2019-09-17
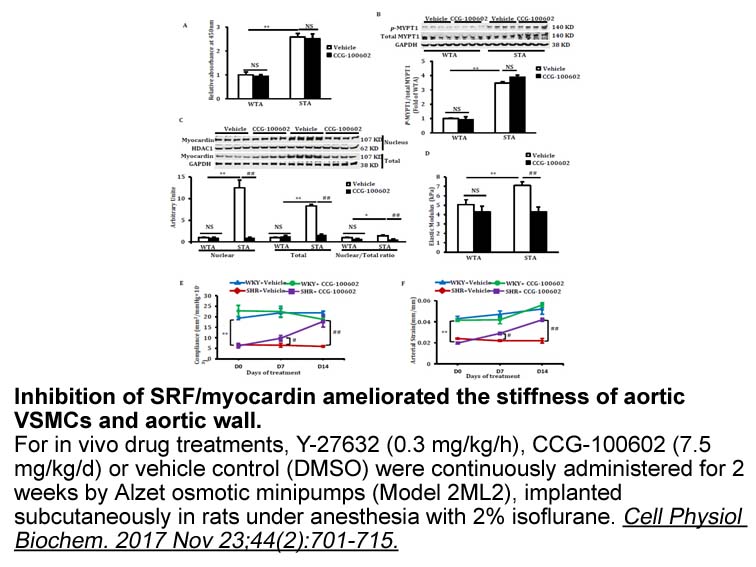
DGKs bind and regulate other signalling proteins Based on the evidence noted above, DGKs achieve specificity of function through a combination of post-translational modifications, the availability of cofactors, and through the availability and access to substrate DAG. DGKs appear to achieve an ad
-
Acknowledgment This work was supported by the Swedish Resear
2019-09-17
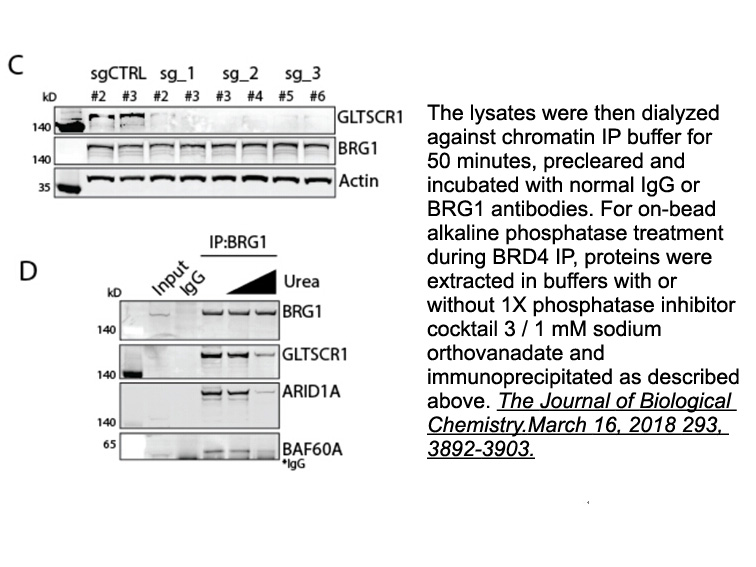
Acknowledgment This work was supported by the Swedish Research Council, the Knut and Alice Wallenberg Foundation, the Göran Gustafsson Foundation, the Swedish Pain Relief Foundation, the Torsten Söderbergs Stiftelse, the Swedish Children\'s Cancer Foundation, and the Swedish Cancer Society. The
-
Gene amplification PCR amplifications were performed using g
2019-09-17
Gene amplification. PCR amplifications were performed using genomic DNA from the different organisms studied as template and two degenerate oligonucleotides (PP1 and PP2) as primersPP1 corresponded to the sense codons of the amino herpes simplex virus 1 motif AGGI(A/S)EM, and PP2 to the antisense s
-
br Introduction The allergic response is
2019-09-17
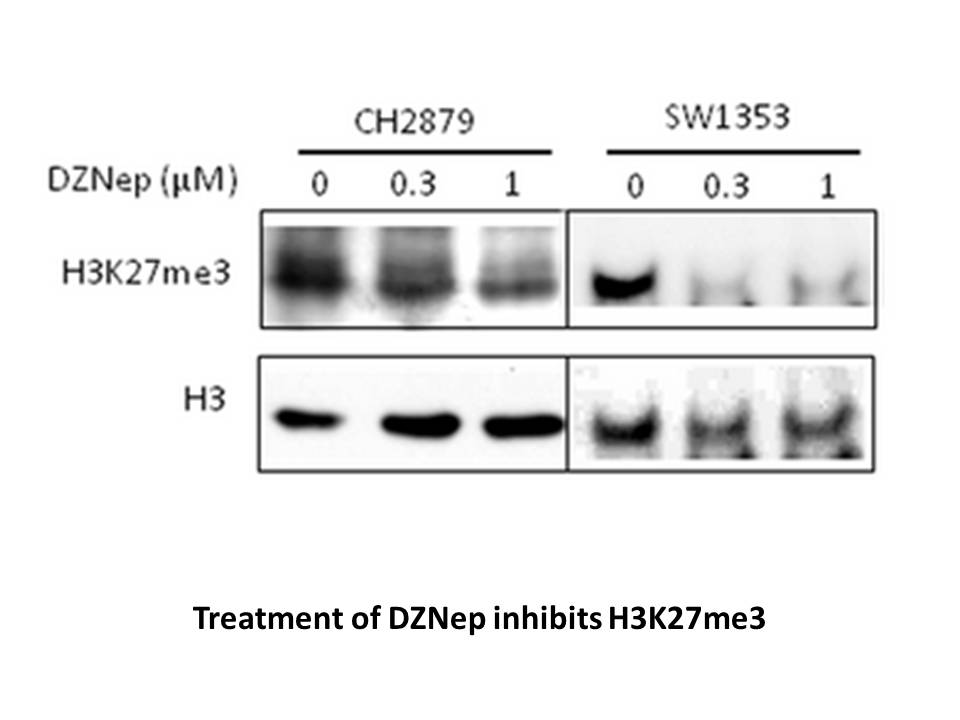
Introduction The allergic response is a complex process involving the interaction of several mediators; among these, cysteinyl leukotrienes (CysLTs) represent one of the most important actors in the pathogenesis of airway allergic diseases such as allergic rhinitis and asthma [1]. Pharmacological
-
Introduction Safety critical systems are defined as those sy
2019-09-17
Introduction Safety-critical systems are defined as those systems where failure could result in the loss of life or injury to people, damage to equipment or to the environment [9]. Safety standards, e.g., [41], [17], [18], [6], [38], expect appropriate safety analysis tasks to be integrated within
-
By using the C elegans Matrisome Annotator tool we found
2019-09-17

By using the C. elegans Matrisome Annotator tool, we found substantial enrichment for matrisome Sulfasalazine in these data sets. Thus, re-analyzing -omic datasets with the C. elegans Matrisome Annotator tool may be useful to generate novel hypotheses about the role of the C. elegans matrisome for
-
Through the process of alternative splicing the IKZF
2019-09-17
Through the process of alternative splicing, the IKZF1 gene is capable of encoding a large number of Ikaros isoforms (Molnar et al., 1996). Some of these isoforms were shown to have distinct functions (Li et al., 2011, Ronni et al., 2007). Ikaros protein contains four zinc fingers at the N-terminus
-
br Introduction Al Awqati and his colleagues using R methyli
2019-09-17
Introduction Al Awqati and his colleagues using R(+)-methylindazone, R(+)-[(6,7-dichloro-2-cyclopentyl-2,3-dihydro-2-methyl-1-oxo-1H-inden-5-yl)-oxy] acetic QS 11 (IAA-94) (Landry et al., 1987) isolated the first chloride intracellular ion channel (CLIC) proteins (p64) from bovine kidney cortex
15847 records 978/1057 page Previous Next First page 上5页 976977978979980 下5页 Last page